Os hormônios vegetais, também chamados fitormônios, são responsáveis pelo controle de processos vitais como germinação, crescimento, desenvolvimento, floração e frutificação das plantas. Assim como ocorre com os nossos hormônios, os vegetais também produzem substâncias capazes de regular todo o seu metabolismo.
Como ocorre nos animais, hormônios vegetais atuam em áreas específicas nas plantas. Cada um deles possui ação sobre determinadas células, tecidos ou órgãos, e possuem uma natureza química diversificada.
São muitos os hormônios vegetais e eles podem sofrer alguma variedade conforme a espécie, mas dentre os principais deles, podemos citar:
- As auxinas
- As giberelinas
- A citocininas
- O etileno gasoso
- O ácido abscísico
Auxinas
As auxinas, principais hormônios das plantas, são produzidas a partir da região apical do caule das plantas, mas também nas folhas mais jovens e nas sementes durante o seu desenvolvimento. A distribuição desse hormônio ocorre de maneira polarizada – do topo para o restante do corpo do vegetal.
A principal atividade das auxinas está relacionada ao crescimento, ao atuar sobre a parede celular, provocando sua elongação ou distensão. O hormônio explica a capacidade de manter indefinidamente o crescimento dos vegetais e, nas espécies que crescem mais rapidamente, possuem maior concentração. Contudo, a depender dessa concentração, as auxinas também podem ter o efeito inverso – inibir o crescimento.
Da mesma forma que hormônios humanos, a concentração e região de atuação das auxinas pode levar a efeitos diferentes nas plantas. Atualmente são conhecidas três auxinas naturais, das quais se destaca o ácido indolacético (IAA), produzido nos ápices dos caules em sementes, em folhas jovens, em flores, frutos e grãos de pólen.
Ação das auxinas
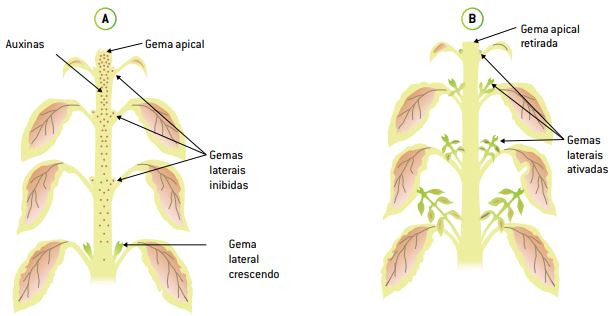
Dominância apical: as auxinas, além de promoverem o processo de distensão celular, quando distribuídas caule abaixo, também inibem o desenvolvimento das gemas laterais, localizadas nas axilas das folhas, que ficam em estado de dormência. Em outras palavras, o hormônio cria uma dominância do eixo central da planta e do caule em relação aos ramos que dele emergem. O hormônio produzido em concentração pelo meristema do caule principal “barra” o excessivo crescimento dos ramos laterais.
Formação de frutos: nas plantas angiospermas, após o processo de fecundação, o embrião presente no interior da semente produz auxinas, que atuam sobre as paredes das células do ovário da flor, promovendo seu desenvolvimento e sua transformação em frutos. Os frutos chamados “verdadeiros” são sempre produzidos conforme esse processo.
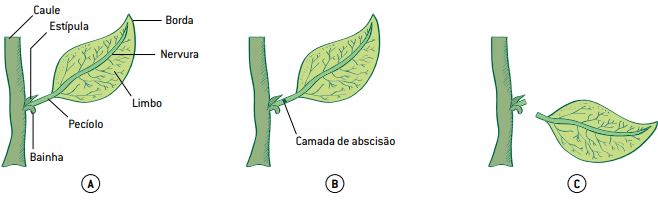
Queda das folhas: a queda das folhas ou abscisão foliar pode ser controlada pela variação na produção de auxinas e do hormônio gasoso etileno, que veremos mais adiante. Quando a produção da auxina cai e a do etileno aumento, as folhas aceleram a sua queda – o que ocorre em especial com plantas que enfrentam temperaturas sazonais mais frias.
Formação de raízes adventícias: há plantas que brotam a partir de mudas, pequenos fragmentos do caule ou mesmo folhas, sem precisar necessariamente de sementes – exemplo disso é a mandioca. Nesses casos, a auxina produzida no caule ou folha induz o segmento da planta a criar raízes adventícias e buscar nutrientes.
Giberelinas
As giberelinas são hormônios vegetais produzidos nos mesmos locais em que são produzidas as auxinas, ou seja, nas gemas apicais, nas folhas jovens e nas sementes em desenvolvimento. As giberelinas também são hormônios de crescimento vegetais. Junto com as auxinas, estimulam o crescimento da planta, promovendo o alongamento das células.
Mesmo em plantas que não produzem ou têm baixa concentração de giberelinas, a aplicação de sintéticos pode promover o crescimento da planta. Além do crescimento, as giberelinas promovem a quebra da dormência de gemas e sementes.
Sementes em dormência impedem a germinação e o desenvolvimento, mas há utilidade nisso em algumas circunstâncias. O teor de giberelina aumenta e a dormência é quebrada – algo que é geralmente disparado pela presença de água. Nesse processo, a giberelina estimula a digestão do amido armazenado no endosperma, fornecendo os nutrientes orgânicos necessários para o desenvolvimento do embrião da planta. As giberelinas também têm um papel fundamental na formação de frutos e na floração.
Citocininas
As citocininas, como a cinetina e a zeatina por exemplo, são substâncias produzidas na região do ápice da raiz e transportadas para partes mais altas do vegetal por meio do xilema a todas as outras partes da planta. Esses hormônios também promovem o crescimento vegetal, acelerando o processo mitótico das células e aumentando seu número no vegetal.
Assim como as giberelinas, as citocininas estão relacionadas com a floração, com o desenvolvimento de gemas laterais e com o retardo da senescência (envelhecimento) da planta. A verdade é que auxinas, giberelinas e citocininas possuem um papel conjunto de regulação no crescimento da planta e na sua reprodução e germinação.
Etileno
O etileno é um hormônio gasoso, produzido por praticamente todos os órgãos das plantas, exceto pelas sementes. Como mostrado anteriormente, em maior concentração e na redução das auxinas, promove a abscisão foliar (queda das folhas), mas também é responsável por acelerar o processo de amadurecimento dos frutos.
Frutos em contêineres fechados são evitados exatamente por essa razão. Se não estão em câmaras frias, os frutos acumulam alta concentração de gás etileno nos contêineres e aceleram seu amadurecimento. Atmosferas pobres em oxigênio também são usadas no transporte de frutos, pois têm o poder de reduzir a taxa de sintetização de etileno.
Ácido abscísico
O ácido abscísico é um hormônio inibidor do crescimento das plantas. Além disso, é responsável pela indução da dormência em sementes e gemas caulinares. Enquanto outros hormônios respondem pela quebra da dormência, o ácido abscísico é responsável por introduzir as sementes nesse estado. Sua concentração tende a aumentar quando as probabilidades de germinação por conta do ambiente externo são pequenas.
O ácido abscísico também está relacionado com o fechamento dos estômatos quando o suprimento de água diminui de forma brusca. Nessa circunstância, a concentração de ácido abscísico aumenta muito nas folhas, o que faz as células-guarda eliminarem potássio, fechando os estômatos. Isso diminui a perda de água pela planta.
Referências:
- MOREIRA, Catarina. Auxinas. Revista de Ciência Elementar, v. 3, n. 4, 2015.
- JORDÁN, Miguel; CASARETTO, José. Hormonas y reguladores del crecimiento: auxinas, giberelinas y citocininas. Squeo, F, A., & Cardemil, L.(eds.). Fisiología Vegetal, p. 1-28, 2006.
- PICOLOTTO, Luciano; BIANCHI, Valmor João; FACHINELLO, José Carlos. Ação de giberelinas e citocininas na germinação de sementes de pessegueiro. Scientia Agraria, v. 8, n. 3, p. 225-232, 2007.
Por: Carlos Artur Matos